
PAL-01: Onde estão os elos perdidos?
Alegação:
Não existem elos perdidos. Onde estão? Ninguém os achou. Tudo o que vemos são lacunas entre os fósseis, e animais surgindo de repente.
Resposta:
“Elo perdido” é um termo popular, e não é utilizado em ciência. O termo correto é fóssil transicional. Isso porque de fato todos os seres vivos que procriaram e já morreram são “elos perdidos” entre um ser vivo e outro.
E existem muitos transicionais, e todos os dias surgem mais. A desculpa para alguém dizer que não existem transicionais é o total desconhecimento deles. E desconhecer uma evidência não significa que ela não exista.
Esses são alguns exemplos de transicionais entre espécie e gênero:
· Ancestralidade humana ( PAL 02) Existem muitos fósseis ancestrais humanos. São cerca de vinte espécies, com centenas de fósseis cada um. E as diferenças vão ocorrendo aos poucos, desde o Australopithecus afarensis até o Homo sapiens.
· Os chifres dos titanotheres (mamíferos extintos do Cenozóico) surgem nas camadas geológicas em tamanhos cada vez maiores, desde o zero. Outros atributos do pescoço e cabeça também evoluíram. Essas adaptações são análogas ao das ovelhas (Stanley 1974).
· Uma seqüência transicional fóssil gradual conecta o foraminífero Globigerinoides trilobus e o Orbulina universa (Pearson et al. 1997). As evidências mostram características sendo adquiridas ao longo do tempo e podem ser vistas nos leitos de todos os oceanos tropicais. Várias morfoespécies intermediárias conectam as duas espécies, que podem ser vistas na figura em Lindsay (1997).
· O registro fóssil mostra transicionais entre os trilobitas Phacops (Eldredge 1972; 1974; Strapple 1978).
· Foranimíferos planctônicos (Malmgren et al. 1984). Esse é um exemplo de gradualismo pontuado. Um registro fóssil de dez milhões de anos mostra longos períodos de êxtase evolutiva, junto com outros períodos de mudanças morfológicas relativamente rápidas, mas ainda assim graduais.
· Fósseis de diatomáceas Rhizosolenia são bastante comuns e mostram mudanças ao longo de quase dois milhões de anos, que incluem um caso de especiação (Miller 1999, 44-45).
· Moluscos do Lago Turkana (Miller 1999, 44-45).
· Ostracodes marinhos do Cenozóico (Cronin 1985).
· O primata do Eoceno do gênero Cantius (Gingerich 1976, 1980, 1983).
· Ostras do gênero Chesapecten mostram mudanças em suas conchas ao longo de 13 milhões de anos (Pojeta and Springer 2001; Ward and Blackwelder 1975).
· Ostras do gênero Gryphaea se tornam maiores e mais largas, porém mais finas e chatas durante o início do Jurássico (Hallam 1968).
Esses são alguns transicionais entre famílias, ordens e classes:
· Ancestralidade humana (PAL 02). O Australopithecus, apesar de suas pernas e pélvis indicarem que ele andava em pé, possuía estrutura óssea no antebraço, provavelmente vestigial, que indica que ele um dia andou se apoiando sobre as mãos (Richmond and Strait 2000).
· Transicionais entre invertebrados e vertebrados ( PAL 03)
· Transicionais entre dinos e aves ( PAL 06).
· Haasiophis terrasanctus é uma cobra primitiva com membros bem desenvolvidos, mostrando um relacionamento com outros ancestrais com patas (Tchernov et al. 2000). Pachyrhachis é outra cobra com patas que é relacionada com a Haasiophis (Caldwell and Lee 1997).
· As mandíbulas dos mosassauros são intermediárias entre cobras e lagartos. Possuíam a mandíbula inferior como a das cobras, flexível e capaz de se esticar. Mas, diferente das cobras, a mandíbula superior não era flexível. Outras características do crânio também são intermediários entre cobras e lagartos primitivos (Caldwell and Lee 1997; Lee et al. 1999; Tchernov et al. 2000).
· Transicionais entre mamíferos e baleias ( PAL 07).
· Transicionais entre peixes e tetrápodes ( PAL 04)
· Transicionais entre condilartos (uma espécie de mamífero terrestre) e os manatis atuais (Domning 2001a, 2001b).
· Runcaria, uma planta do Médio Devoniano, foi o precusor das plantas com semente. Possuía todas as qualidades das sementes, exceto um envoltório sólido para elas, e um sistema para guiar o pólen até a semente (Gerrienne et al. 2004).
· A abelha Melittosphex burmensis, preservada em ambar do Cretácio Inferior, possui características primitivas transicionais entre vespas e abelhas (Poinar and Danforth 2006).
Esses são alguns transicionais entre reino e filo:
· Os fósseis do Cambriano Halkiera e Wiwaxia possuem características que os ligam um ao outro e aos filos modernos Mollusca, Brachiopoda, e Annelida (Conway Morris 1998, 185-195).
· Os fósseis Anomalocaris e Opabinia, do Cambriano e Pré-cambriano, são transicionais entre artrópodes e lobópodes.
· Um ancestral dos equinodermos foi encontrado, que é um intermediário entre os equinodermos modernos e os deuterostomas (Shu et al. 2004).
· Recentemente descoberto, um fóssil transicional une sozinho três grandes filos: anelídeos, moluscos e braquiópodes. Consultar em: http://arstechnica.com/journals/science.ars/2007/3/1/7278
Referências:
Caldwell, M. W. and M. S. Y. Lee, 1997. A snake with legs from the marine Cretaceous of the Middle East. Nature 386: 705-709.
Conway Morris, Simon, 1998. The Crucible of Creation, Oxford University Press.
Cronin, T. M., 1985. Speciation and stasis in marine ostracoda: climatic modulation of evolution. Science 227: 60-63.
Domning, Daryl P., 2001a. The earliest known fully quadupedal sirenian. Nature 413: 625-627.
Domning, Daryl P., 2001b. New “intermediate form” ties seacows firmly to land. Reports of the National Center for Science Education 21(5-6): 38-42.
Eldredge, Niles, 1972. Systematics and evolution of Phacops rana (Green, 1832) and Phacops iowensis Delo, 1935 (Trilobita) from the Middle Devonian of North America.
Bulletin of the American Museum of Natural History 147(2): 45-114.
Eldredge, Niles, 1974. Stability, diversity, and speciation in Paleozoic epeiric seas. Journal of Paleontology 48(3): 540-548.
Gerrienne, P. et al. 2004. Runcaria, a Middle Devonian seed plant precursor. Science 306: 856-858.
Gingerich, P. D., 1976. Paleontology and phylogeny: Patterns of evolution of the species level in early Tertiary mammals. American Journal of Science 276(1): 1-28.
Gingerich, P. D., 1980. Evolutionary patterns in early Cenozoic mammals. Annual Review of Earth and Planetary Sciences 8: 407-424.
Gingerich, P. D., 1983. Evidence for evolution from the vertebrate fossil record. Journal of Geological Education 31: 140-144.
Hallam, A., 1968. Morphology, palaeoecology and evolution of the genus Gryphaea in the British Lias. Philosophical Transactions of the Royal Society of London B 254: 91-128.
Lee, Michael S. Y., Gorden L. Bell Jr. and Michael W. Caldwell, 1999. The origin of snake feeding. Nature 400: 655-659.
Lewin, R., 1981. No gap here in the fossil record. Science 214: 645-646.
Lindsay, Don, 1997. A smooth fossil transition: Orbulina, a foram. http://www.don-lindsay-archive.org/creation/orbulina.html
Malmgren, B. A., W. A. Berggren and G. P. Lohmann, 1984. Species formation through punctuated gradualism in planktonic foraminifera. Science 225: 317-319.
Miller, Kenneth R., 1999. Finding Darwin’s God. New York: HarperCollins.
Pearson, P. N., N. J. Shackleton and M. A. Hall. 1997. Stable isotopic evidence for the sympatric divergence of Globigerinoides trilobus and Orbulina universa (planktonic foraminifera). Journal of the Geological Society, London 154: 295-302.
Poinar, G. O. Jr. and B. N. Danforth. 2006. A fossil bee from Early Cretaceous Burmese amber. Science 314: 614.
Richmond B. G. and D. S. Strait, 2000. Evidence that humans evolved from a knuckle-walking ancestor. Nature 404: 382-385. See also Collard, M. and L. C. Aiello, 2000. From forelimbs to two legs. Nature 404: 339-340.
Shu, D.-G. et al., 2004. Ancestral echinoderms from the Chengjiang deposits of China. Nature 430: 422-428.
Stanley, Steven M., 1974. Relative growth of the titanothere horn: A new approach to an old problem. Evolution 28: 447-457.
Strapple, R. R., 1978. Tracing three trilobites. Earth Science 31(4): 149-152.
Tchernov, E. et al., 2000. A fossil snake with limbs. Science 287: 2010-2012. See also Greene, H. W. and D. Cundall, 2000. Limbless tetrapods and snakes with legs. Science 287: 1939-1941.
Ward, L. W. and B. W. Blackwelder, 1975. Chesapecten, A new genus of Pectinidae (Mollusca: Bivalvia) from the Miocene and Pliocene of eastern North America. U.S. Geological Survey Professional Paper 861.
Mais informações:
Hunt, Kathleen. 1994-1997. Transitional vertebrate fossils. FAQ. http://www.talkorigins.org/faqs/faq-transitional.html
Miller, Keith B. n.d. Taxonomy, transitional forms, and the fossil record. http://www.asa3.org/ASA/resources/Miller.html
Patterson, Bob. 2002. Transitional fossil species and modes of speciation. http://www.origins.tv/darwin/transitionals.htm
Thompson, Tim. 1999. On creation science and transitional fossils. http://www.tim-thompson.com/trans-fossils.html
Cohn, Martin J. and Cheryll Tickle. 1999. Developmental basis of limblessness and axial patterning in snakes. Nature 399: 474-479. (technical)
Cuffey, Clifford A. 2001. The fossil record: Evolution or “scientific creation”. http://www.gcssepm.org/special/cuffey_00.htm or http://www.nogs.org/cuffeyart.html
Elsberry, Wesley R. 1995. Transitional fossil challenge. http://www.rtis.com/nat/user/elsberry/evobio/evc/argresp/tranform.html
Godfrey, L. R. 1983. Creationism and gaps in the fossil record. In: Godfrey, L. R. (ed.), Scientists Confront Creationism, New York: W. W. Norton, pp. 193-218.
Morton, Glenn R. 2000. Phylum level evolution. http://home.entouch.net/dmd/cambevol.htm
Pojeta, John Jr. and Dale A. Springer. 2001. Evolution and the Fossil Record, Alexandria, VA: American Geological Institute, http://www.agiweb.org/news/spot_06apr01_evolutionbk.htm , http://www.agiweb.org/news/evolution.pdf, pg. 2.
Strahler, Arthur N. 1987. Science and Earth History, Buffalo, NY: Prometheus Books, pp. 398-400.
Zimmer, Carl. 2000. In search of vertebrate origins: Beyond brain and bone. Science 287: 1576-1579.
Ainda mais informações:
Golfinhos com resquício de patas traseiras
Fóssil descoberto no RS é considerado marco evolutivo
Pássaros exibem seleção natural em tempo real
Transicional entre vespas e abelhas
Ainda muito mais transicionais
Depois que os criacionistas lerem todos os trabalhos desses cientistas (e muitos outros), e os refutarem no âmbito da ciência, ou seja, com pesquisas científicas, teses e publicações com peer review, aí sim eles podem dizer que não existem transicionais. Antes disso é pura cegueira e teimosia.
PAL-02: Hominídeos são 100% homens ou 100% macacos
Alegação:
Os fósseis hominídeos são na verdade totalmente humanos ou totalmente macacos.
Resposta:
Não é verdade. Há uma fina transição entre os fósseis de hominídeos, onde não é possível estabelecer claramente uma linha onde o fóssil deixa de ser humano. Tanto que os próprios criacionistas divergem entre eles sobre o que eles consideram humano e o que eles consideram macaco (Foley 2002).
O fato é que a maioria dos criacionistas que dizem isso sequer seguraram um crânio de algum hominídeo na vida. Ou seja, falam apenas por convicção pessoal, aquilo que eles acham ou esperam que seja, sem se importar se é verdade ou não.
Para exemplificar isso, exemplos de transicionais humanos nos mostram que:
· Australopithecus afarensis, que viveu entre 3,9 a 3,0 milhões de anos atrás, possui o crânio parecido com os chimpanzés atuais, mas com os dentes mais humanos. A maioria (provavelmente todos) dos criacionistas diriam que era um macaco, apesar de ser bípede;
· Australopithecus africanus (3 a 2 milhões de anos), com um cérebro entre 420 a 500 centímetros cúbicos, um pouco maior que o A. afarensis, e com dentes ainda mais humanos;
· Homo habilis (2,4 a 1,5 milhões de anos), parecido com os australopithecus, mas usava ferramentas e tinha um cérebro maior (650 centímetros cúbicos em média) e um rosto menos projetado, mais humano;
· Homo erectus (1,8 a 0,3 milhões de anos); cérebro entre 900 cm³ nos primeiros H. erectus e 1.100 cm³ nos últimos. (O cérebro moderno possui em média 1.350 cm³;
· O Homo sapiens do Pleistoceno era “morfologicamente e cronologicamente intermediário entre fósseis arcaicos africanos e os anatomicamente mais modernos fósseis do Pleistoceno superior” (White et al. 2003, 742).
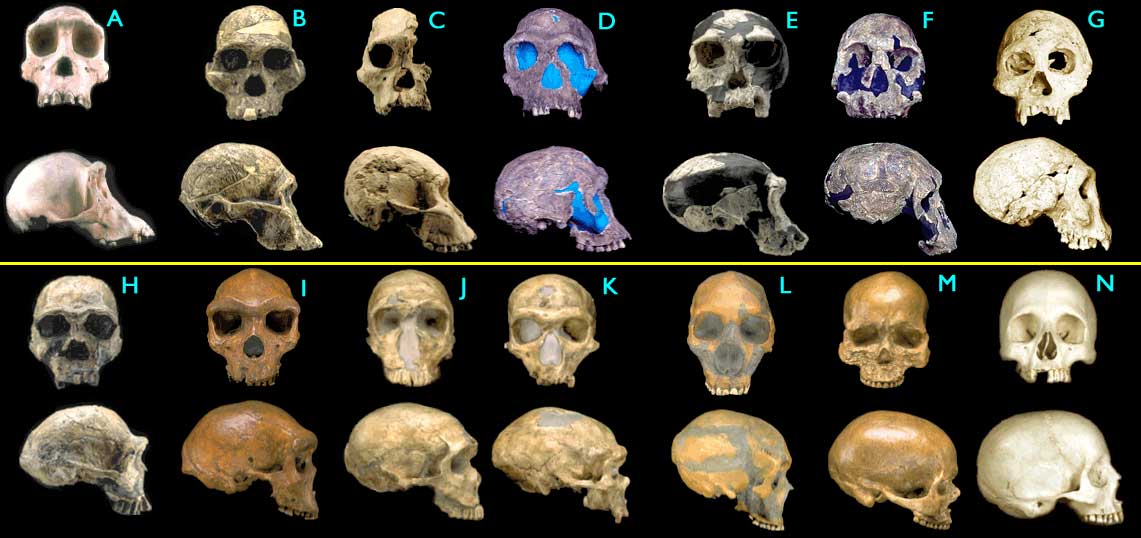
A imagem neste link abaixo mostra claramente a evolução humana ao longo do tempo:
http://anthropologynet.files.wordpress.com/2007/06/fossil-hominid-skulls.jpg
(A) Pan troglodytes, chimpanzé moderno. Está ali apenas para comparação. Não faz parte da evolução humana.
(B) Australopithecus africanus
(C) Australopithecus africanus
(D) Homo habilis
(E) Homo habilis
(F) Homo rudolfensis
(G) Homo erectus
(H) Homo ergaster
(I) Homo heidelbergensis
(J) Homo sapiens neanderthalensis (não faz parte da evolução humana)
(K) Homo sapiens neanderthalensis
(L) Homo sapiens neanderthalensis
(M) Homo sapiens sapiens, Cro-Magnon I
(N) Homo sapiens sapiens, homem moderno
Referências:
Avarello, R., A. Pedicini, A. Caiulo, O. Zuffardi, M. Fraccaro, 1992. Evidence for an ancestral alphoid domain on the long arm of human chromosome 2. Hum Genet 89(2): 247-249.
Bermudez de Castro, J. M. et al., 1997. A hominid from the Lower Pleistocene of Atapuerca, Spain: Possible ancestor to Neandertals and modern humans. Science 276: 1392-1395.
Foley, Jim, 1996-2003. (see above)
Foley, Jim, 2002. Comparison of all skulls, http://www.talkorigins.org/faqs/homs/compare.html
IJdo, J. W., A. Baldini, D. C. Ward, S. T. Reeders and R. A. Wells, 1991. Origin of human chromosome 2: an ancestral telomere-telomere fusion. Proceedings of the National Academy of Science USA 88(20): 9051-9055.http://www.pnas.org/cgi/reprint/88/20/9051.pdf
Max, Edward E., 2003. Plagiarized errors and molecular genetics. http://www.talkorigins.org/faqs/molgen/
Taylor, D. M. 2003. Alignment of Chimp_rp43-42n4 against human chromosome 15. http://www-personal.umich.edu/~lilyth/erv/ See also Taylor, D. M. 2003 (Jun 3). Re: Evolutionary Misconceptions on Evolution. http://www.google.com/groups?as_umsgid=75200cbc.0306031846.50b2bda5%40posting.google.com
White, Tim D. et al., 2003. Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423: 742-747.
Yunis, J. J. and O. Prakash, 1982. The origin of man: a chromosomal pictorial legacy. Science 215: 1525-1530.
Foley, Jim. 1996-2004. Fossil hominids: The evidence for human evolution. http://www.talkorigins.org/faqs/homs/
Drews, Carl, 2002. Transitional fossils of hominid skulls. http://www.theistic-evolution.com/transitional.html
Johanson, D. C., and B. Edgar, 1996. From Lucy to Language. New York: Simon and Schuster.
Leakey, M. and A. Walker, 1997. Early hominid fossils from Africa. Scientific American 276(6) (June): 74-79.
Tattersall, Ian, 1995. The Fossil Trail. New York: Oxford.
http://www.talkorigins.org/faqs/comdesc/hominids.html
PAL-03: Fósseis entre vertebrados e invertebrados?
Alegação:
Não existem fósseis transicionais entre vertebrados e invertebrados.
Resposta:
Os seguintes fósseis do Cambriano são transicionais entre vertebrados e invertebrados:
· Pikaia, um cordado invertebrado primordial. Inicialmente foi interpretado como um verme segmentado, até que novas análises mostraram que ele possuía notocorda;
· Yunnanozoon, outro cordado primordial;
· Haikouella, um cordado similar ao Yunnanozoon, mas com características adicionais, como um coração e um cérebro relativamente maiores (Chen et al. 1999);
· Conodontes possuíam dentes ósseos, mas o resto do seu corpo era mole. Eles também possuíam notocorda (Briggs et al. 1983; Sansom et al. 1992);
· Cathaymyrus diadexus, o mais antigo cordado conhecido, 535 milhões de anos (Shu et al. 1996);
· Myllokunmingia e Haikouichthys, dois vertebrados primitivos que ainda não tinham uma cabeça, estrutura óssea e dentes plenamente definidos. Eles diferem de invertebrados mais antigos por terem um arranjo dos músculos segmentados em zigue-zague (Monastersky 1999).
Existem alguns animais vivos hoje que são “quase-vertebrados”, e podem ser considerados formas intermediárias, como o anfioxo e as lampreias. O estudo desses animais pode revelar muito sobre a origem dos vertebrados.
Referências:
Briggs, D. E. G., E. N. K. Clarkson and R. J. Aldridge, 1983. The conodont animal. Lethaia 16: 1-14.
Chen, J.-Y., D.-Y. Huang and C.-W. Li, 1999. An early Cambrian craniate-like chordate. Nature 402: 518-522.http://www.nature.com/cgi-taf/DynaPage.taf?file=/nature/journal/v402/n6761/full/402518a0_r.html
Monastersky, Richard, 1999. Waking up to the dawn of vertebrates. Science News 156: 292.http://www.sciencenews.org/sn_arc99/11_6_99/fob1.htm
Sansom, I. J., M. P. Smith, H. A. Armstrong and M. M. Smith, 1992. Presence of the earliest vertebrate hard tissues in conodonts. Science 256: 1308-1311.
Shu, D.-G., S. Conway Morris and X.-L. Zhang, 1996. A Pikaia-like chordate from the Lower Cambrian of China. Nature 384: 157-158.
Mais informações:
Speer, B. R. 2000. Introduction to the Deuterostomia. http://www.ucmp.berkeley.edu/phyla/deuterostomia.html
# Waggoner, Ben. 1996. Introduction to the Cephalochordata. http://www.ucmp.berkeley.edu/chordata/cephalo.html
PAL-04: Fósseis entre peixes e anfíbios?
Alegação:
Não existem transicionais entre peixes e tetrápodes.
Resposta:
Existem vários bons transicionais:
· A maioria dos peixes possuem aberturas nasais externas anteriores e posteriores. Nos tetrápodes, a abertura nasal posterior é substituída pelo coano (choana), orifícios das fossas nasais que se abrem na rinofaringe, e que nenhum peixe possui. Kenichthys, um fóssil de 395 milhões de anos da China, é um intermediário exato entre os dois (Zhu and Ahlberg 2004).
· Um fóssil mostra oito dedos ósseos em sua barbatana frontal, mostrando que os dedos surgiram antes dos tetrápodes saírem da água (Daeschler and Shubin 1998).
· Um úmero do Devoniano tem características que mostram que pertenceu a um tetrápode aquático que conseguia se impulsionar para frente com seus membros superiores, mas não era capaz de mover os inferiores para frente e para trás para caminhar (Shubin et al. 2004).
· Acanthostega, outro fóssil do Devoniano, com cerca de 60 cm, e que provavelmente viveu em rios (Coates 1996). Tinha polidactia em seus membros, mas sem ligamentos dos pulsos ou tornozelos (Coates e Clack 1990). Era predominantemente, se não totalmente, aquático. Tinha guelras como os peixes (Coates e Clack 1991), e seus membros e coluna não conseguiam suportar muito peso.
· Ichthyostega, mais um tetrápode do Devoniano, tinha cerca de 1,5 m e provavelmente era anfíbio. Tinha sete dedos nas patas traseiras (patas dianteriras são desconhecidas). Seus membros e coluna eram mais robustos que oAcanthostega. Tinha espinha parecida com peixe em sua cauda, mas eram menores que as do Acanthostega. Seu crânio apresenta várias características de peixes primitivos, mas sem guelras (Murphy 2002).
· Tulerpeton, de depósitos estuários mais ou menos da mesma época que o Acanthostega e o Ichthyostega, tinha seis dedos nos membros frontais e sete nos traseiros. Seus ombros eram mais robustos que o Acanthostega, sugerindo que eram um pouco menos aquáticos, e seu crânio era mais parecido com anfíbios do Carbonífero Superior do que com o Acanthostega ou o Ichthyostega.
Referências:
Coates, M. I. 1996. The Devonian tetrapod Acanthostega gunnari Jarvik: postcranial anatomy, basal tetrapod interrelationships and patterns of skeletal evolution. Transactions of the Royal Society of Edinburgh: Earth Sciences 87: 363-421.
Coates, M. I. and J. A. Clack, 1990. Polydactyly in the earliest known tetrapod limbs. Nature 347: 66-69.
Coates, M. I. and J. A. Clack, 1991. Fish-like gills and breathing in the earliest known tetrapod. Nature 352: 234-236.
Daeschler, Edward B. and Neil Shubin, 1998. Fish with fingers? Nature 391: 133.
Murphy, Dennis C., 2002. Devonian times: Ichthyostega stensioei. http://www.mdgekko.com/devonian/Order/re-ichthyostega.html
Shubin, N. H., E. B. Daeschler and M. I. Coates, 2004. The early evolution of the tetrapod humerus. Science 304: 90-93. See also: Clack, J. A., 2004. From fins to fingers. Science 304: 57-58.
Zhu, Min and Per E. Ahlberg, 2004. The origin of the internal nostril of tetrapods. Nature 432: 94-97. See also: Janvier, Philippe, 2004. Wandering nostrils. Nature 432: 23-24.
Mais informações:
http://www.orkut.com/CommMsgs.aspx?cmm=184959&tid=2453223899302182571
PAL-05: Fósseis entre anfíbios e répteis?
Alegação:
Não há fósseis entre anfíbios e répteis.
Resposta:
A principal diferença entre os anfíbios (tetrápodes primitivos) e os répteis (amniotas) é a presença do âmnion, que não fossiliza. Por isso é extremamente difícil diferenciar um do outro, até porque suas estruturas ósseas são muito parecidas.
No entanto essa incerteza não nos dá o direito de dizer categoricamente que não existem transicionais entre eles. Existem vários fósseis de criaturas do Permiano, alguns são amniotas primitivos, outros provavelmente não são.
Esses são alguns exemplos de transicionais entre anfíbios e répteis:
· Reptilomorfos, como o Eogyrinus e o Diadectes
· Archaeothyris, um amniota primitivo do Carbonífero basal
· Batrachosauria, também do Carbonífero
Mais informações:
http://www.palaeos.com/Vertebrates/Units/190Reptilomorpha/190.100.html
PAL-06: Fósseis entre dinos e aves?
Alegação:
Não há fósseis entre dinossauros e aves.
Resposta:
Muitos fósseis de aves foram descobertos nas últimas décadas, vários deles revelaram serem intermediários entre dinossauros (como os Allosauros) e aves modernas:
· Sinosauropteryx prima, um dinossauro coberto por penas primitivas, mas estruturalmente similar aos dinossauros sem penas Ornitholestes e Compsognathus (Chen et al. 1998; Currie and Chen 2001).;
· Ornitomimossauros, therizinossauros e oviraptorssauros. O oviraptorosauro Caudipteryx tinha o corpo coberto com tufos de penas (Ji et al. 1998). Também possuía penas o therizinosauro chamado Beipiaosaurus (Xu et al. 1999a). Várias outras características de aves são encontradas nesses dinossauros, como crânio e vértebras altamente pneumatizados, e longas asas. Oviraptórides também tinham ovos similares aos das aves, e inclusive os chocavam (Clark et al. 1999);
· Deinonychosauros (troodontides e dromaeosauros). Esses são os dinossauros mais próximos das aves. Sinovenator, o mais primitivo troodontide, é bastante similar ao Archaeopteryx (Xu et al. 2002). Byronosauro, outro troodontide, possuía dentes praticamente idênticos às aves primitivas (Makovicky et al. 2003). Microraptor, o mais primitivo dromaeosaur, é também o mais parecido com ave; espécimes foram encontrados com irrefutáveis penas em suas asas, pernas e cauda (Hwang et al. 2002; Xu et al. 2003). Sinornithosauro também era coberto por uma variedade de penas e um crânio mais parecido com aves do que os dromaeosauros mais antigos (Xu, Wang, and Wu 1999; Xu and Wu 2001; Xu et al. 2001);
· Protarchaeopteryx, alvarezsaurídeos, Yixianosaurus e Avimimus. Esses são dinossauros de locais incertos, cada um potencialmente mais próximo das aves do que os deinonychosauros. Protarchaeopteryx tem penas na cauda, e asas/patas alongadas (Ji et al. 1998). Yixianosaurus possui uma distinta plumagem nas asas/patas proporcional às aves (Xu and Wang 2003). Alvarezsaurídeos (Chiappe et al. 2002) e Avimimus (Vickers-Rich et al. 2002) possuem outras características de aves;
· Archaeopteryx. Esse famoso fóssil está classificado com ave, mas ele possui mais características de dinossauros do que alguns dos citados acima.(Paul 2002; Maryanska et al. 2002);
· Shenzhouraptor (Zhou and Zhang 2002), Rahonavis (Forster et al. 1998), Yandangornis e Jixiangornis. Todas essas aves eram ligeiramente mais avançados que o Archaeopteryx, especialmente nas vértebras, esterno e ossos das asas;
· Sapeornis (Zhou and Zhang 2003), Omnivoropteryx e confuciusornithids (ex., Confuciusornis e Changchengornis; Chiappe et al. 1999). Esses são as primeiras aves a possuírem grandes ossos formados por vértebras da cauda fusionados (chamados pigostilos);
· Enantiornitinos, incluindo pelo menos dezenove espécies de aves primitivas, como o Sinornis (Sereno and Rao 1992; Sereno et al. 2002), Gobipteryx (Chiappe et al. 2001), e Protopteryx (Zhang and Zhou 2000). Várias novas características de aves aparecem, incluindo doze ou menos vértebras dorsais, fúrcula estreita em forma de V (o chamado osso da sorte), e redução nos ossos digitais das asas;
· Patagopteryx, Apsaravis, e yanornithids (Chiappe 2002; Clarke and Norell 2002). Mais características de aves aparecem nesses grupos, incluindo mudanças nas vertebras desenvolvimento de quilha, expansão do osso esterno, onde se prendem os músculos das asas;~
· Hesperornis, Ichthyornis, Gansus, e Limenavis. Essas aves são quase tão avançadas quanto aves modernas. Novas características incluem perda de quase todos os dentes, e mudanças nos ossos das pernas;
· Finalmente, aves modernas.
Referências:
Chen, P., Z. Dong and S. Zhen, 1998. An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature 391: 147-152.
Chiappe, L. M., 2002. Osteology of the flightless Patagopteryx deferrariisi from the Late Cretaceous of Patagonia (Argentina). In Chiappe and Witmer, pp. 281-316.
Chiappe, L. M. and L. M. Witmer (eds.), Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: Univ. of California Press.
Chiappe, L. M., M. A. Norell and J. M. Clark, 2001. A new skull of Gobipteryx minuta (Aves: Enantiornithes) from the Cretaceous of the Gobi Desert. American Museum Novitates 3346: 1-15. http://diglib1.amnh.org/novitates/i0003-0082-346-01-0001.pdf
Chiappe, L. M., M. A. Norell and J. M. Clark, 2002. The Cretaceous, short-armed Alvarezsauridae. In: Chiappe and Witmer, pp. 87-120.
Chiappe, L. M., S. Ji, Q. Ji and M. A. Norell, 1999. Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of northeastern China. Bulletin of the American Museum of Natural History 242: 1-89.http://diglib1.amnh.org/bulletins/i0003-0090-242-01-0001.pdf
Clark, J. M., M. A. Norell and L. M. Chiappe, 1999. An oviraptorid skeleton from the Late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avianlike brooding position over an oviraptorid nest. American Museum Novitates 3265: 1-36.
Clarke, J. A. and M. A. Norell, 2002. The morphology and phylogenetic position of Apsaravis ukhaana from the late Cretaceous of Mongolia. American Museum Novitates 3387: 1-46. http://diglib1.amnh.org/novitates/i0003-0082-3387-01-0001.pdf
Currie, P. J. and P. Chen, 2001. Anatomy of Sinosauropteryx prima from Liaoning, northeastern China. Canadian Journal of Earth Sciences 38: 1705-1727.
Forster, C. A., S. D. Sampson, L. M. Chiappe and D. W. Krause, 1998. The theropod ancestry of birds: New evidence from the Late Cretaceous of Madagascar. Science 279: 1915-1919.
Hwang, S. H., M. A. Norell, Ji Q. and Gao K., 2002. New specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from northeastern China. American Museum Novitates 3381: 1-44.http://research.amnh.org/users/sunny/hwang.et.al.2002.pdf
Ji, Q., P. Currie, M. A. Norell and S-A. Ji, 1998. Two feathered dinosaurs from northeastern China. Nature 393: 753-761.
Makovicky, P. J., M. A. Norell, J. M. Clark and T. Rowe, 2003. Osteology and relationships of Byronosaurus jaffei (Theropoda: Troodontidae). American Museum Novitates 3402, 1-32. http://diglib1.amnh.org/novitates/i0003-0082-3402-01-0001.pdf
Maryanska, T., H. Osmólska and M. Wolsan, 2002. Avialan status for oviraptorosauria. Acta Palaeontologica Polonica 47(1): 97-116. http://app.pan.pl/acta47/app47-097.pdf
Paul, Gregory S., 2002. Dinosaurs of the Air. Baltimore: Johns Hopkins University Press.
Sereno, P. C. and C. Rao, 1992. Early evolution of avian flight and perching: New evidence from the Lower Creates of China. Science 255: 845-848.
Sereno, P. C., C. Rao and J. Li, 2002. Sinornis santensis (Aves: Enantiornithes) from the Early Cretaceous of Northeastern China. In: Chiappe and Witmer, pp. 184-208.
Vickers-Rich, P., L. M. Chiappe and S. Kurzanov, 2002. The enigmatic birdlike dinosaur Avimimus portentosus. In: Chiappe and Witmer, pp. 65-86.
Xu, X. and X. Wang, 2003. A new maniraptorian dinosaur from the Early Cretaceous Yixian Formation of Western Liaoning. Vertebrate Palasiatica 41(3): 195-202.
Pojeta, John Jr. and Springer, Dale A., 2001. Evolution and the Fossil Record, American Geological Institute, Alexandria, VA. http://www.agiweb.org/news/spot_06apr01_evolutionbk.htm ,http://www.agiweb.org/news/evolution.pdf .
Prum, Richard O. and Alan H. Brush, 2003. Which came first, the feather or the bird? Scientific American 288(3) (Mar.): 84-93.
Wang, Justin, 1998. Scientists flock to explore China’s ‘site of the century’. Science 279: 1626-1627.
PAL-07: Fósseis entre mamíferos e cetáceos?
Alegação:
Não existem fósseis entre mamíferos e baleias.
Resposta:
Existem vários transicionais que provam que as baleias e os golfinhos se originaram de mamíferos terrestres. Babinski (2003) e Zimmer (1998) apresentam várias figuras para os seguintes:
· Pakicetus inachus: do final do Eoceno primário (Gingerich et al. 1983; Thewissen and Hussain 1993);
· Ambulocetus natans: Do início ao médio Eoceno. Tinha pequenos membros frontais e patas traseiras adaptadas ao nado. Sua coluna ondulava para cima e para baixo, facilitando o nado. Aparentemente podia andar em terra tão bem quando nadir (Thewissen et al. 1994);
· Indocetus ramani: início do médio Eoceno (Gingerich et al. 1993);
· Dorudon: o cetáceo dominante do final do Eocene. Os seus pequenos membros traseiros não serviam para locomoção;
· Basilosaurus: Médio Eoceno. Uma baleia aquática completa com pernas plenamente estruturadas (Gingerich et al. 1990);
· Uma baleia primitive com seu respire muito mais à frente e algumas outras estruturas encontradas em animais terrestres, mas não em baleias mais recentes (Stricherz 1998).
Os parentes vivos mais próximos das baleias são os hipopótamos. Um grupo fóssil conhecido como anthracotheres liga os hipopótamos às baleias (Boisserie et al. 2005).
A evolução das baleias é nítida também no crânio desses animais. Os mais primitivos possuíam ouvidos ótimos para escutar fora d’água, mas ruins dentro dela. À medida que eles iam se adaptando para os ambientes marinhos, seus ouvidos iam melhorando para escutar dentro d’água, e piorando para escutar fora dela, até que atualmente os ouvidos das baleias são excelentes para dentro d’água, e ruins para escutar fora.
Referências:
Boisserie, Jean-Renaud, Fabrice Lihoreau and Michel Brunet. 2005. The position of Hippopotamidae within Cetartiodactyla. Proceedings of the National Academy of Science USA 102(5): 1537-1541.
Gingerich, P. D. et al., 1983. Origin of whales in epicontinental remnant seas: New evidence from the Early Eocene of Pakistan. Science 220: 403-406.
Gingerich, P. D., B. H. Smith, and E. L. Simons, 1990. Hind limb of Eocene Basilosaurus: Evidence of feet in whales. Science 249: 154-157.
Gingerich, P. D. et al., 1993. Partial skeletons of Indocetus ramani [Mammalia, Cetacea] from the Lower Middle Eocene Domanda Shale in the Sulaiman Range of Punjab [Pakistan]. Contributions from the Museum of Paleontology of the University of Michigan 28: 393-416.
Gingerich, P. D. et al., 1994. New whale from the Eocene of Pakistan and the origin of cetacean swimming. Nature 368: 844-847.
Gingerich, P. D. et al. 2001. Origin of whales from early artiodactyls: Hands and feet of Eocene Protocetidae from Pakistan. Science 293: 2239-2242. See also: Rose, K. D. 2001. The ancestry of whales. Science 293: 2216-2217.
Thewissen, J. G. M. and S. T. Hussain, 1993. Origin of underwater hearing in whales. Nature 361: 444-445.
Thewissen, J. G. M., S. T. Hussain and M. Arif, 1994. Fossil evidence for the origin of aquatic locomotion in archaeocete whales. Science 263: 210-212. See also Berta, A., 1994. What is a whale? Science 263: 180-181.
Stricherz, Vince, 1998 (10 Oct.). Burke displays fossil of toothless whale. http://depts.washington.edu/uweek/archives/1998.10.OCT_29/_article2.html See also http://more.abcnews.go.com/sections/science/DailyNews/baleen980916.html
Babinski, E. T., 2003. Cetacean evolution (whales, dolphins, porpoises) http://www.edwardtbabinski.us/babinski/whale_evolution.html
Sutera, Raymond, 2001. The origin of whales and the power of independent evidence. Reports of the National Center for Science Education 20(5): 33-41. http://www.talkorigins.org/features/whales/
Gould, S. J. 1995. Hooking leviathan by its past. In: Dinosaur in a Haystack. New York: Harmony Books, pp. 359-376.
Pojeta, John Jr. and Dale A. Springer. 2001. Evolution and the Fossil Record, American Geological Institute, Alexandria, VA. http://www.agiweb.org/news/spot_06apr01_evolutionbk.htm ,http://www.agiweb.org/news/evolution.pdf .
Thewissen, J. G. M. (ed.). 1998. The Emergence of Whales: evolutionary patterns in the origin of Cetacea. New York: Plenum. (technical)
Thewissen, J. G. M., S. I. Madar, and S. T. Hussain. 1998. Whale ankles and evolutionary relationships. Nature 395: 452. See also Wong, K., 1999 (Jan.). Cetacean creation. Scientific American 280(1): 26,30.
Thewissen, J. G. M. and E. M. Williams. 2002. The early radiations of Cetacea (Mammalia): Evolutionary pattern and developmental correlations. Annual Review of Ecology and Systematics 33: 73-90. (technical)
Zimmer, Carl. 1995. Back to the sea. Discover 16(1) (Jan.): 82-84.
Zimmer, Carl. 1998. At the Water’s Edge. New York: Touchstone, ch. 6-10.
PAL-08: O Homem de Piltdown foi uma fraude
Alegação:
Tem mais coisa feia!!! Alguns dos ossos do passado, famosos por serem considerados de espécies transicionais, não são mais vistos dessa maneira nem pelos evolucionistas. O homem de Piltdown, uma referencia nos livros de ciência e museus durane anos acabou desmascarado como fraude.
Resposta:
Uma das vedetes dos argumentos criacionistas, por algum motivo eles citam fraudes que ocorreram décadas atrás como se elas invalidassem todos os fósseis que já foram encontrados até hoje.
É claro que não, fraudes ocorrem em qualquer área de conhecimento. Mas foi o método científico que permitiu descobrir que se tratava de uma montagem de um crânio humano com o maxilar de um chimpanzé.
Só isso seria suficiente para derrubar o pobre argumento de Piltdown, mas há mais coisas a serem ditas.
Piltdown, antes de ser desmascarado pela ciência como fraude, não era aceito com unanimidade pela comunidade científica. Na verdade, ele foi uma pedra no sapato, pois ele não se encaixava na teoria evolucionária. É claro, era um híbrido inventado com partes de ossos modernos.
Ele foi aceito por muitos apenas por ser um achado fabuloso para a época, o que é uma fraqueza humana, a qual todos nós estamos sujeitos.
A fraude durou por quarenta anos, gerando muita polêmica, pois seus grandes descobridores, Charles Dawson e Arthur Smith Woodward, proibiram o acesso a análises científicas, deixando o crânio fora do alcance de todos, dentro do Museu Britânico. Somente quando eles morreram, Piltdown foi desmascarado como a fraude que era. Vale ressaltar, desmascarado por cientistas, graças ao método científico.
E não nos esqueçamos que os perpetradores da fraude eram criacionistas.
Descobrir um erro não invalida o método científico, muito pelo contrário, ele apenas o fortalece. Assim sabemos que podemos confiar na ciência, pois ela se corrige, se aprimora, e se fiscaliza.
Interessante notar, após Piltdown ser descoberto como forjado, nenhum cientista jamais o citou como verdadeiro. No entanto, muitos criacionistas continuamente citam fraudes, não importando quantas vezes elas já foram desmascaradas, como as pegadas de humanos e dinossauros em Paluxy, esqueletos de gigantes, descoberta da Arca de Noé, Darwin se arrepender no leito de morte, citações de cientistas fora de contexto, etc.
Referências:
Harter, Richard, 1996. Piltdown Man: The bogus bones caper.
PAL-09: O Homem de Nebraska foi uma fraude
Alegação:
De um dente, montaram a mandíbula. Da mandíbula, montaram o crânio. Do crânio, montaram o esqueleto. Do esqueleto, fizeram pele, cabelo e até a sua namorada ou esposa (agachada no desenho). Fizeram a famosa exposição sobre a evolução em Dayton, Tennessee, chamada de Scopes Trial, quando o Homem de Nebraska foi apresentado como prova incontestável da evolução. Quando William Jennings Bryan protestou contra os argumentos apresentados e pela insuficiência, riram-se dele ridicularizando-o. Em 1927 descobriram a fraude: O dente era de um porco chamado Peccary.
Fonte: http://www.baptistlink.com/creationists/pilt.htm
Resposta:
A história não é bem assim.
O dente não foi considerado como evidência por muitos cientistas. A maioria deles na verdade eram céticos quanto à descoberta, diferente de como os criacionistas querem fazer parecer.
O próprio Henry Fairfield Osborn, paleontólogo que descreveu o dente, estava em dúvidas o dente pertencia aos hominídeos ou a outra espécie de primata:
“Até estarmos mais seguros quanto a arcada dentária, ou partes do crânio, ou do esqueleto, não podemos estar certos se o Hesperopithecus pertence aos Simiidae ou aos Hominidae.” (Osborn 1922)
O tal desenho que os criacionistas gostam de usar para ridicularizar a evolução, foi feito por uma revista de cunho popular, tipo Superinteressante ou Veja, e não foi nem jamais teve intenção de ser cientificamente correta. Tal desenho jamais foi publicado em trabalhos científicos.
Até Osborn se espantou ao ver o desenho, e disse que:
“Tal desenho ou ‘reconstrução’ seria com certeza apenas proveniente da imaginação humana, sem valor científico nenhum, e com certeza impreciso” (Wolf and Mellett 1985)
A alegação de que o Homem de Nebraska foi usado no Scopes “Monkey” Trial é mentirosa. Os criacionistas que mostrem as transcrições do julgamento se quiserem provar isso.
O Homem de Nebraska foi um exemplo de ciência funcionando bem. Uma descoberta intrigante foi encontrada, podendo ter várias implicações. A descoberta foi anunciada e vários outros experts a analisaram. Cientistas de início eram céticos. Mais evidências foram encontradas, e a conclusão foi que a interpretação inicial estava errada. Finalmente, a retratação foi publicada.
Referências:
Foley, Jim, 2001. Creationist arguments: Nebraska Man. http://www.talkorigins.org/faqs/homs/a_nebraska.html
Mellett, James S. and John Wolf, 1985. The role of “Nebraska man” in the creation-evolution debate. Creation/Evolution 16: 31-43. http://www.talkorigins.org/faqs/homs/wolfmellett.html
Gould, S. J., 1991. An essay on a pig roast. In Bully for Brontosaurus, pp. 432-47. New York: W.W.Norton.
Osborn H.F. (1922): Hesperopithecus, the anthropoid primate of western Nebraska. Nature, 110:281-3.
PAL-10: O Homem de Neandertal sofria de raquitismo
Alegações:
O Homem de Neanderthal: foi reconstituído a partir de um crânio quase completo descoberto em 1848 e um esqueleto parcial em 1856. Muitos estudiosos dizem que o Neanderthal era tão humano quanto qualquer um de nós. As diferenças do esqueleto são atribuídas ao fato de pertencer a um homem velho que sofria de raquitismo. Esse detalhe foi comprovado com novos achados fósseis, pois os Neanderthais sepultavam seus mortos
Os darwinistas argumentaram que o homem neandertal era uma criatura semelhante ao macaco, enquanto muitos críticos de Darwin (como o grande anatomista Rudokph Virchow) argumentaram que os Neandertais eram humanos em todo aspecto, apesar de que alguns aparentavam sofrer de raquitismo ou artrite.
Fonte: http://www.pibvp.org.br/index.php?name=News&file=article&sid=289
Resposta:
É óbvio que quando os primeiros neandertais foram descobertos, os cientistas não sabiam ao certo onde eles se encaixavam na evolução humana. Seríamos nós descendentes diretos deles? Ou são eles de uma linhagem paralela? Ou seriam eles apenas pessoas com doenças, como raquitismo ou artrite?
Essas dúvidas até eram normais e compreensíveis há mais de um século atrás, porém hoje, com mais de 300 espécimes de neandertais descobertos, sabemos que eles eram na verdade de uma linhagem paralela à nossa. Temos até evidências genéticas disso.
Eles eram nossos primos, uma outra espécie de ser humano. É possível até que nossos antepassados tenham se encontrado com eles.
É verdade que alguns deles podem ter sofrido de raquitismo, ou de artrite, porém nem de longe a maioria, muito menos todos, sofreram dessas doenças.
E outra, raquitismo e artrite não transformam uma pessoa em um neandertal.
Pessoas que sofrem de raquitismo têm alta deficiência em cálcio, e seus ossos são tão mais fracos que até o peso do próprio corpo pode quebrá-los
Já os neandertais possuíam ossos 50% mais grossos que os dos humanos comuns, e possuíam porte atlético e robusto.
Dizer que eles eram idênticos aos seres humanos atuais é torcer os fatos. Sua estrutura óssea é completamente distinta dos seres humanos.
Mas isso não faz deles macacos. Em muitos aspectos eles eram parecidos com os humanos; andavam sobre os dois pés, tinham hábitos de comunidades, etc. Mas não eram idênticos a nós. Eram uma outra espécie de ser humano.
Quem sabe nossos ancestrais tenham até lutado com eles por supremacia, e nós fomos os vitoriosos. A jornada da humanidade no nosso planeta é fascinante, é muito triste ver como os criacionistas querem varrer nossa história para baixo do tapete, escondê-la das pessoas, só porque ela desmente suas crenças e seus mitos.
Referências:
Straus, W. L. Jr. and A. J. E. Cave, 1957. Pathology and the posture of Neanderthal man. Quarterly Review of Biology 32(4): 348-363.
Foley, Jim. 2002. Creationist arguments: Neandertals. http://www.talkorigins.org/faqs/homs/a_neands.html
PAL-11: Australopithecus são totalmente macacos
Alegação:
Australopithecus são macacos completos, próximos do chimpanzé. Solly Zuckerman e Charles Oxnard, evolucionistas renomados, mostraram que eles não andavam em pé.
Resposta:
Australopithecus africanus e robustus são muito mais parecidos com humanos do que com macacos. A. afarensis fica mais ou menos no meio termo entre os dois, talvez pendendo mais para os primatas. A. ramidus é mais simiesco.
As alegações de que eles são totalmente primatas vem de apenas dois artigos já desacreditados pela ciência. O próprio Oxnard considerava o Australopithecus um ancestral humano, sem problemas (Groves 1999). Há vastas evidências de que aquelas espécies eram plenamente bípedes e tinham ainda outras características humanóides. Essas evidências são ignoradas, distorcidas, ou simplesmente negadas pelos criacionistas (Foley 1997).
Referências:
Foley, Jim. 1997. Creationist arguments: Australopithecines. http://www.talkorigins.org/faqs/homs/a_piths.html
Groves, Colin. 1999. Getting desperate. http://www.talkorigins.org/faqs/homs/desperate_cg.html
PAL-12: Archaeopteryx era apenas um pássaro
Alegação:
Archaeopteryx era um pássaro completo. Possuía penas e asas como um pássaro comum.
Resposta:
Archaeopteryx está definido como um pássaro. No entanto, possuía muito mais características de dinossauros do que de aves.
Suas principais características de aves eram:
· Longas narinas externas;
· Quadrato e quadratojugal (ossos da mandíbula) não unidos;
· Ossos do palato que possuíam três extensões;
· Grande canal lateral no topo da parte traseira das vértebras;
· Outras características de aves do Archaeopteryx são encontradas em vários outros dinossauros, como penas, fúrcula (osso da sorte), e púbis alongada e direcionada para trás;
· As principais características de dinossauros são as seguintes;
· Dentes nos ossos do maxilar e premaxilar;
· Garras nas asas, com três dedos;
· Longa cauda com ossos;
· Abertura nasal bem na frente, separada dos olhos por uma grande abertura preorbital;
· Costelas finas, não unidas ou articuladas com o externo;
· Sacro que ocupa seis vértebras;
· Metacarpos livres (exceto pelo terceiro);
· Pulso da mão flexível;
· Pélvis e rótula do fêmur no formato dos arcossauros em muitos detalhes;
· Ossos da pélvis não fundidos.
E mais de 100 outras diferenças das aves (Chiappe 2002; Norell and Clarke 2001).
Referências:
Nedin, Chris. 1999. All about Archaeopteryx. http://www.talkorigins.org/faqs/archaeopteryx/info.html
Chiappe, L. M. 2002. Basal bird phylogeny. In: Chiappe and Witmer, pp. 448-472.
Chiappe, L. M. and L. M. Witmer (eds.). 2002. Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: Univ. of California Press.
Elzanowski, A. 2002. Archaeopterygidae (Upper Jurassic of Germany). In: Chiappe and Witmer, pp. 129-159.
Mayr, Gerald, Burkhard Pohl, and D. Stefan Peters. 2005. A well-preserved Archaeopteryx specimen with theropod features. Science 310: 1483-1486.
Middleton, K. M. 2002. Evolution of the perching foot in theropods. Journal of Vertebrate Paleontology 22: 88A.
Norell, M. A. and J. A. Clarke. 2001. Fossil that fills a critical gap in avian evolution. Nature 409: 181-184.
PAL-13: Archaeopteryx foi uma fraude
Alegação:
As impressões de penas do Archaeopteryx de Londres foram forjadas. Existem evidências de que:
· As impressões das penas aparecem em apenas uma placa, e não na outra oposta.
· Fissuras do tamanho de fios de cabelo atravessando ambos ossos e penas podem ter sido formados por movimentos nas placas depois que o cimento estava no lugar.
· As penas parecem “carimbos duplicados”.
· A textura da superfície é diferente entre as áreas com penas e as sem.
· Pequenas “bolhas” aparecem que nem sempre se encaixam na placa oposta.
· Sob o microscópio, a rocha parece diferente a parte fóssil e não-fóssil do espécime.
· Materiais desconhecidos aparecem na malha do fóssil.
· Análise química de raios-X mostra diferenças químicas que incluem silicone, sulfetos e cloro na área fóssil que não estão presentes na área não-fóssil.
Esses pontos indicam que as penas foram impressas por alguém que usou algum tipo de molde. Sem as penas, Archaeopteryx seria identificado como o dinossauro Compsognathus, que não seria transicional.
Fonte:
Watkins, R. S., F. Hoyle, N. C. Wickramasinghe, J. Watkins, R. Rabilizirov, and L. M. Spetner, 1985a. Archaeopteryx — a photographic study. British Journal of Photography 132: 264-266.
Watkins, R. S. et al., 1985b. Archaeopteryx — a further comment. British Journal of Photography 132: 358-359,367.
Watkins, R. S. et al., 1985c. Archaeopteryx — further evidence. British Journal of Photography 132: 468-470.
Hoyle, Fred, N. C. Wickramasinghe and R. S. Watkins, 1985. Archaeopteryx: Problems arise — and a motive. British Journal of Photography 132(6516): 693-695,703.
Hoyle, Fred and C. Wickramasinghe, 1986. Archaeopteryx, The Primordial Bird, Christopher Davis, London.
Spetner, L. M., F. Hoyle, N. C. Wickramasinghe and M. Magaritz, 1988. Archaeopteryx — more evidence for a forgery. British Journal of Photography 135: 14-17.
Resposta:
Existem outros nove fósseis de Archaeopteryx descobertos em lugares e épocas diferentes, todos muito bem documentados. Pelo menos seis deles possuem claras e indiscutíveis penas (Charig 1986; Wellnhofer 1993; Mayr et al. 2005). No exemplar Maxburg, as penas continuam até os ossos, acabando com qualquer possibilidade de fraude (Charig 1986). Além disso, vários outros dinossauros com penas foram encontrados.
Pequenas fraturas, infiltradas por calcita, se estendem entre penas e ossos, mostrando que ambos vieram da mesma fonte. As fissuras também se encaixam perfeitamente nas placas opostas (Charig 1986). Essas fraturas são invisíveis a olho nu, então um fraudador do século XIX sequer saberia de sua existência, muito menos conseguiria replicá-las.
O efeito de “carimbo duplo” na placa oposta se deve ao método de fossilização. Bactérias crescem debaixo das penas, degradando-as, fazendo os sedimentos abaixo litificarem, e assim preservando uma impressão mais forte das penas. Quando as penas decaíram, os sedimentos acima pressionaram para baixo criando um molde na superfície de baixo (Davis and Briggs 1995). Evidência desse processo, incluindo bactérias litificadas, são visíveis sob microscópio de alta resolução, e não poderiam de modo algum serem forjadas.
A diferença de textura entre as áreas com e sem penas se deve ao próprio corpo do animal (Charig 1986).
As “bolhas” são irregularidades naturais. Não há sequer uma que não tenha uma depressão correspondente na placa oposta. As duas metades se encaixam bem exceto onde a superfície foi destruída para a preparação subseqüente (Charig 1986).
As aparências diferentes nas áreas fóssil e não fóssil se devem a diferentes resoluções usadas nas fotografias do microscópio SEM (Nedin 1997).
Os materiais desconhecidos claramente não estão dentro da malha fóssil (Spetner et al. 1988, Figs. 4b-f). Os grãos de carbonato são apenas poeira.
As diferenças químicas entre a área fóssil e não fóssil são resultado do tratamento de preservação aplicado sobre a área fóssil (Nedin 1997).
Não adianta. Archaeopteryx é um legítimo transicional dino-ave, quer os criacionistas gostem ou não. Podem até tentar, mas não vão conseguir ofuscar essa maravilhosa descoberta, essa prova incrível de que o legado dos dinossauros sobreviveu entre nós, na forma das aves que povoam a terra e os céus…
Referências:
Nedin, Chris, 1997. On Archaeopteryx, astronomers, and forgery. http://www.talkorigins.org/faqs/archaeopteryx/forgery.html
Charig, Alan J. et al. 1986. Archaeopteryx is not a forgery. Science 232: 622-626.
Davis, Paul G. and Derek E. G. Briggs. 1995. Fossilization of feathers. Geology 23(9): 783-786.
Mayr, Gerald, Burkhard Pohl, and D. Stefan Peters. 2005. A well-preserved Archaeopteryx specimen with theropod features. Science 310: 1483-1486.
Spetner, L. M., F. Hoyle, N. C. Wickramasinghe and M. Magaritz. 1988. Archaeopteryx – more evidence for a forgery. British Journal of Photography 135: 14-17.
Swinburne, N. H. M. 1988. The Solnhofen Limestone and the preservation of Archaeopteryx. Trends in Ecology and Evolution 3(10): 274-277.
Wellnhofer, P. 1993. The seventh specimen of Archaeopteryx from the Solnhofen Limestone. Archaeopteryx 11: 1-47.
Majka, Christopher, 1992. Archaeopteryx – is this bird a fraud? New Brunswick Naturalist http://www.chebucto.ns.ca/Environment/NHR/archaeopteryx.html
PAL-14: Archaeoraptor foi uma fraude
Alegação:
Archaeoraptor foi considerado como um transicional entre dinossauros e aves (Sloan 1999), mas foi revelado como uma fraude, um composto com o corpo de ave com a cauda de um dinossauro.
Resposta:
Archaeoraptor não foi uma fraude científica. Ele foi montado por um caçador de fósseis chinês que o descobriu. As partes foram montadas para fazer o fóssil mais valioso para colecionadores, não para pesquisadores. Talvez o seu descobridor sequer sabia que eram de fósseis diferentes (Simons 2000).
O Archaeoraptor foi publicado em revistas populares, não em jornais com peer-review. O autor do artigo principal foi o diretor de arte da revista National Geographic, não um cientista. Ambas Nature e Science rejeitaram os artigos descrevendo-o, alegando suspeitas de que ele foi forjado e ilegalmente contrabandeado (Dalton 2000; Simons 2000).
Procedimentos científicos naturais foram mantidos em todos os seus padrões normais de rigor, até ser revelado como fraude, pela própria ciência, diga-se de passagem.
Vale lembrar que as duas metades do Archaeoraptor (Yanornis martini, o corpo, e Microraptor zhaoianus, a cauda), são fósseis valiosos por si só (Rowe et al. 2001; Xu et al. 2000; Zhou et al. 2002).
Referências:
Dalton, Rex, 2000. Feathers fly over Chinese fossil bird’s legality and authenticity. Nature 403: 689-690.
Rowe, T. et al., 2001. The Archaeoraptor forgery. Nature 410: 539-540.
Sloan, Christopher P., 1999. Feathers for T. Rex? National Geographic 196(5) (Nov.): 98-107.
Xu, Xing, Zhonghe Zhou and Xiaolin Wang, 2000. The smallest known non-avian theropod dinosaur. Nature 408: 705-708.
Zhou, Zhonghe, Julia A. Clarke and Fucheng Zhang, 2002. Archaeoraptor’s better half. Nature 420: 285.
Simons, Lewis M., 2000. Archaeoraptor fossil trail. National Geographic 198(4) (Oct.): 128-132.

{kind=link}
{kind=link}